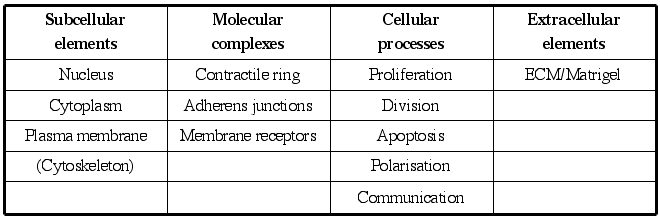
IBCell model structure: Intra-, inter- and extra-cellular elements included in the immersed boundary model of a eukaryotic cell:
Morphological changes in cell undergoing certain life processes:
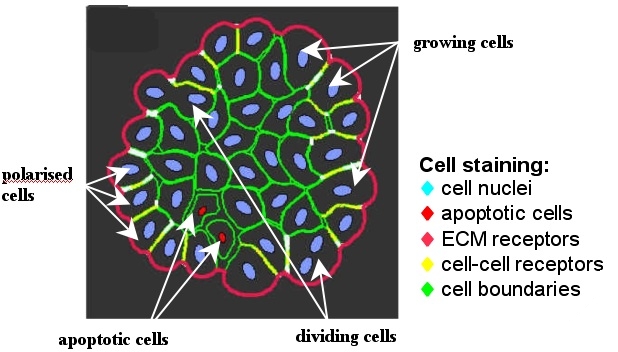
Modeling approaches and computer simulations of individual cell life processes:

Cell growth and division in 3D
The same algorithm as in 2D case is used here, however, the contractile ring is formed by short linear springs acting along the cell equator.

Orientation of the axis of cell division: orientation of cell division depends on the cell shape and its location within the cell cluster.
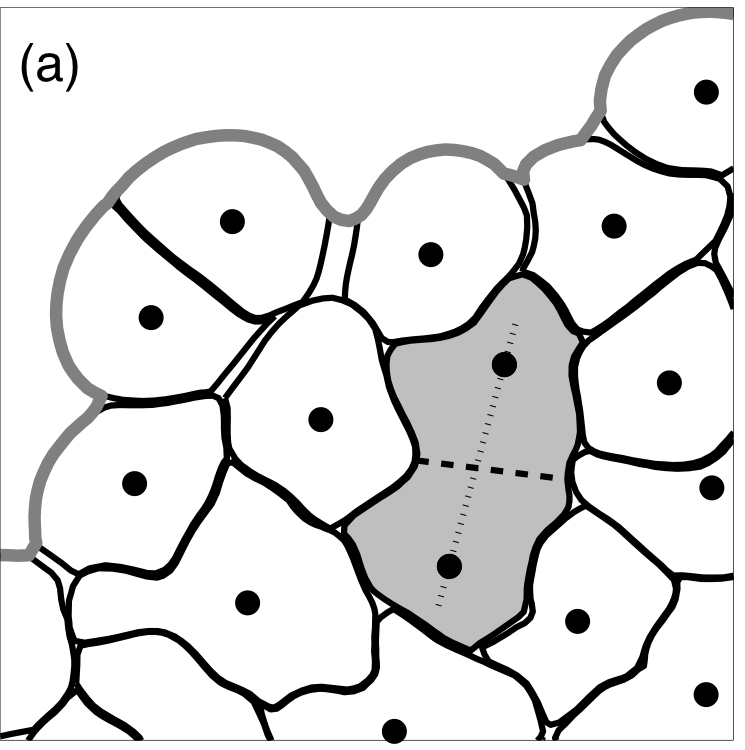
Division in an unpolarised epithelial cell
In a dividing unpolarised cell (a dark grey cell) two new nuclei are located along the cells longest axis (a thin line joining two nuclei) and the contractile ring (a dark dashed line) is placed between them orthogonally to this axis in such a way as to split the cell into two approximately equal parts.
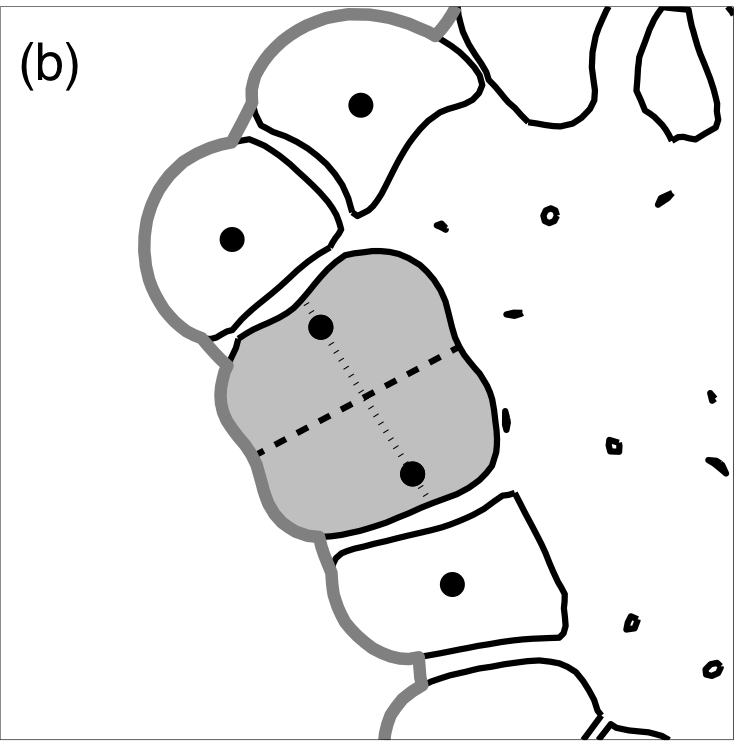
Division in a polarised epithelial cell
The orientation of the axis of cell division in a polarised cell (a dark grey cell) is orthogonal to the part of cell membrane that is in contact with the ECM (a dark dashed line). Two new daughter nuclei are then placed orthogonal to the axis of cell division giving rise to two luminal cells.
Cell apoptotic death:

Cell apoptosis in 3D
An apoptotic cell undergoes very characteristic changes in its morphology, including detachment from the neighbouring cells, shrinkage of the cell volume, collapse of the cytoskeleton and alternations in the cell surface resulting in a bubbling appearance.
Cell epithelial polarization:
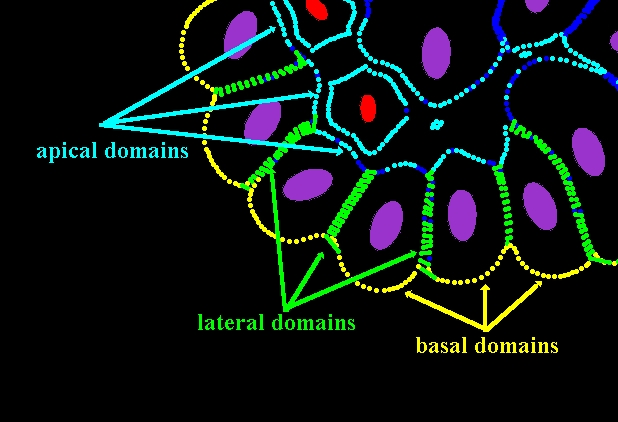
Epithelial cell polarisation in 3D
The mature epithelial cells acquire three different cell membrane domains: an unbounded apical side facing free space inside the epithelium (lumen) and two bounded sides that are in direct contact with either the extracellular matrix (a basal side) or the membranes of other cells (lateral sides). Cell polarity is determined by recognising lateral and basal sides and by developing the apical side and tight junctions along its lateral sides.
Cell chemotactic movement:

Cell-cell interaction during movement:
cell-cell interactions during cell movement
![]()
More detail on particular cell life processes, their numerical implementation within the immersed boundary framework, and computatinal simulatins can be found in:
• K.A. Rejniak, "An immersed boundary framework for modelling the growth of individual cells: an application to the early tumour development", Journal of Theoretical Biology, 2007, 247:186-204. available on: http://dx.doi.org/10.1016/j.jtbi.2007.02.019
• K.A. Rejniak, "Modelling the Development of Complex Tissues using Individual Viscoelastic Cells", chapter IV.3 of the book "Single-Cell-Based Models in Biology and Medicine" edited by A.R.A. Anderson, M.A.J. Chaplain, K.A.Rejniak, Birkhauser-Verlag, Mathematics and Biosciences in Interaction (MBI) series, 2007, ISBN 978-3-7643-8101-1.
• K.A. Rejniak, A.R.A. Anderson, "A computational study of the development of epithelial acini: I. Sufficient conditions for the formation of a hollow structure", Bulletin of Mathematical Biology, 2008, 70(3):677-712. available on: http://dx.doi.org/10.1007/s11538-007-9274-1
![]()